Decompacting Identity: New Imaging Technologies and the Frozen Brain

Introduction

Figure 1. PHerc. Paris. 4, the first successfully read Herculaneum scroll.
Carrie Radomski recently published an essay that compares cryonics to the recovery of the Herculaneum scrolls through the Vesuvius Challenge (Radomski, 2026). It is a fine introduction to the information-theoretic argument for cryonics. While I agree with some of her analysis, I believe the analogy between preserved brains and carbonized scrolls has crucial limitations. Those limitations bear directly on how we should think about information loss, freezing injury, and the prospects of revival for frozen (as opposed to vitrified) patients. They may also shape the future of research into the recoverability of cryopreserved patients.
As she notes in her article, the analogy between cryopreserved brains and the Herculaneum scrolls is not exact. Several differences merit consideration. Crucially, the scrolls “only” changed substrates. Intense heating converted the organic papyrus into a carbon-rich, almost entirely inorganic residue (Figure 1) while preserving enough of its gross and microscopic structure to permit modern imaging and reconstruction. Thus, the problem wasn’t so much information loss as it was the transition to a substrate whose fragility and configuration made it impossible to access them as originally intended—namely, by unrolling them and distinguishing ink from papyrus (Figure 2).

Figure 2. Using the wrong technology to recover information can result in catastrophic, irreversible loss, as seen above. For roughly 250 years, scholars repeatedly damaged scrolls in attempts to read them. As the standard histories note, physical opening often damaged or destroyed the manuscripts. Camillo Paderni’s early method involved slicing scrolls apart. In 1816, Molard and Rochette destroyed an entire scroll while attempting to unroll it. In 1877, a papyrus taken to the Louvre was subjected to mechanical opening and was mostly destroyed, leaving only about one-quarter intact. Reproduced from Ceccarelli et al., 2025.
This is not to minimize the tour de force of the method used to recover their information content: particle-accelerator synchrotrons to capture ultra-high-resolution X-ray data, coupled with machine-learning systems capable of detecting extraordinarily subtle differences in texture and thickness associated with the ink-bearing fibers (Parsons et al., 2023; Ceccarelli et al., 2025). This specific technological approach would have been viewed as unprecedented five years ago, while the foundational idea of non-invasive reading would have seemed like science fiction a decade ago.
A Multiplicity of Injuries
However, the analogy breaks down because injury to preserved human brains is multifactorial, heterogeneous, and highly variable. Some patients will have comparatively little or even no brain injury from their terminal illnesses or from aging-related neurodegeneration, little injury from ischemia, and, compared to uncryoprotected freezing (UCP), much less injury from the preservation technique. Others will have experienced extensive age-related neurodegeneration, ischemic injury, and inadequate or no cryoprotection at all. Currently, it is probably reasonable to characterize most patients as having a combination of these conditions. In such cases, the better analogy may be the image in Figure 2, which resulted from an unsuccessful attempt to read a Herculaneum scroll in the 19th century. Some of this damage is irreversible under our current understanding of physics; stirred ashes no longer contain the information required for reconstruction.
The same is true in biostasis, where brain structure degrades and is lost during prolonged neurodegeneration. Similarly, ischemia and reperfusion injury range from “modest” biophysical changes—such as cellular edema and non-destructive structural alterations that do not preclude the inference of information, such as nuclear chromatin clumping and mitochondrial calcification—to extensive free-radical- and enzymatic-mediated degradation of cell membranes.
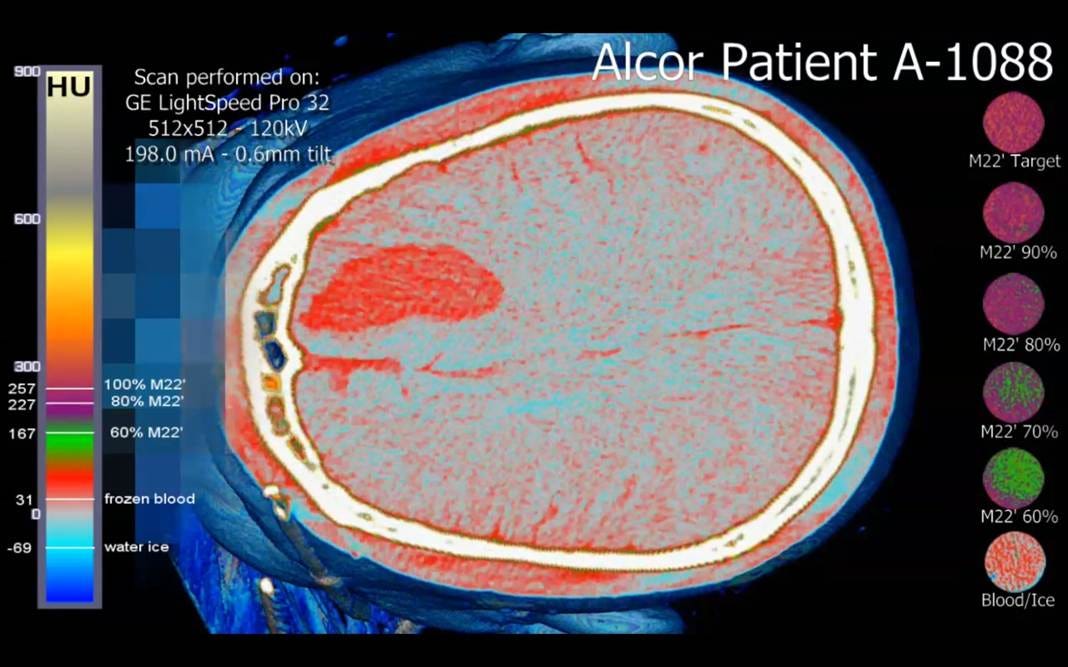
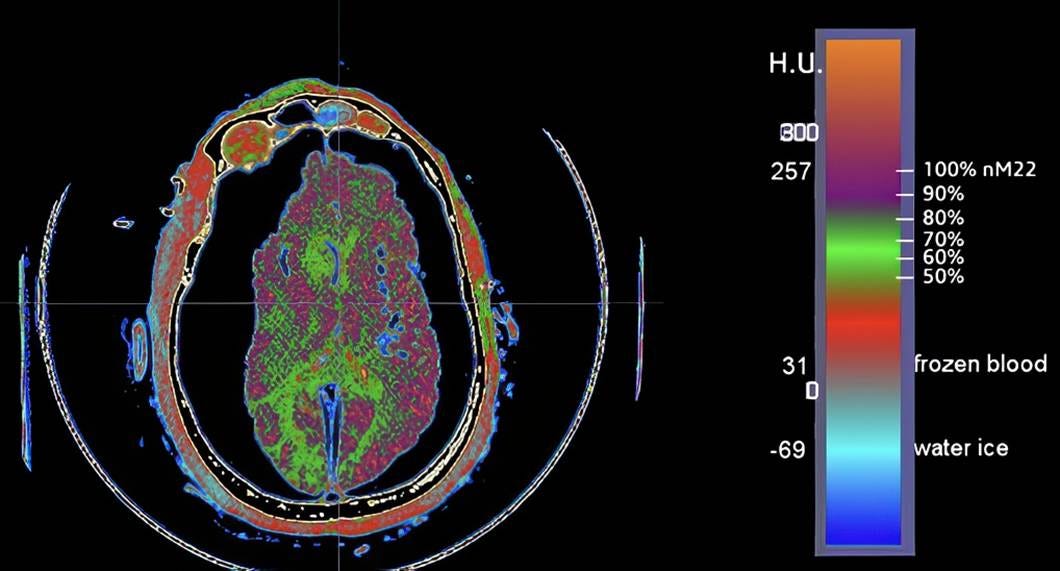
Figure 3. Top: Cryogenic CT scan of a patient who suffered an intracranial bleed in the right prefrontal cortex and was pronounced dead by neurological criteria, precluding cryoprotective perfusion. (Baldwin, 2013). There is extensive ice formation in the cerebral parenchyma (pale blue). Bottom: Cryogenic CT scan of a patient (A-2705) who experienced significant, multimolar cryoprotection throughout the brain, with some areas of inadequate cryoprotectant equilibration resulting in ice formation (green areas). The brain is markedly dehydrated due to osmotic water removal by cryoprotectant agents during perfusion.
Damage from ice formation may range from modest spatial displacement of the connectome to extensive physical remodeling, rendering it impossible to recover the necessary information (Figures 5 and 6). Almost all patients today, regardless of how they have been treated, experience significant regional brain ice formation, and a significant minority undergo extensive ice formation, or “complete” freezing, due to the lack of any meaningful cryoprotection (Figure 3). The latter is particularly concerning because ice formation under these conditions generates enormous hydraulic forces on the tissue trapped between the advancing ice fronts. The tissue is not just “dehydrated” by ice; it is squeezed by a moving, multi-dimensional network of ice that exerts compressive and shear forces across many spatial planes simultaneously (Figures 7 and 8).
Of interest is that, as far as I can determine, cryobiology has not considered the three-dimensional morphology of ice formation in bulk frozen tissues. Studies appear to have been exclusively confined to two-dimensional views using cryomicroscopy and single-slice freeze-substitution light and transmission electron microscopy.
A Look Inside the Frozen-Thawed Brain
Figure 3 illustrates the typical appearance of neuropil in a young, healthy animal during a study conducted by my colleagues and me in 1982-1983. Figures 5 and 6 are also from that study and show what brain ultrastructure looks like in animals subjected to 30 minutes of warm ischemia, followed by ≈ 24 hours of cold ischemia (packed in ice), after which cryoprotective perfusion with 4 M glycerol was carried out, and the animals were frozen to liquid nitrogen temperature (-196 ℃). FIG is an abbreviation for “feline ischemia glycerol.” The micrographs were made after the animals were thawed, the brain tissue was fixed with aldehydes, and the samples were examined by transmission microscopy (TEM). Because the animals could not be perfused with fixative due to unanticipated macroscopic fracturing, the tissues for microscopy had to be sliced and immersion-fixed. This raised concerns that these manipulations may have caused or exacerbated the observed injury. The complete results of that study are available here and provide additional information about the nature and extent of cryoinjury under the conditions used to treat cryonics patients during the “glycerol cryoprotection era.” These results are likely still relevant to today’s patients, in whom cryoprotection is suboptimal and large amounts of ice form, and comparable results have been observed in the brain slice freezing and vitrification work of Pichugin, Fahy & Morrin (2006)
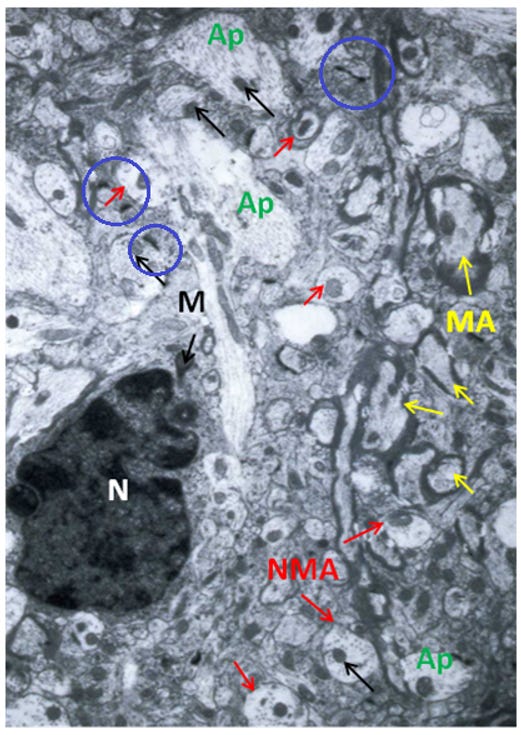
Figure 4: Control, Hippocampus, original magnification 15,000x. This section shows mostly non-myelinated (NMA) axons, with an astrocyte’s nucleus (N) dominating the micrograph’s left side. The axoplasm (Ap) and mitochondria (M), black arrows, exhibit normal density and architecture. Several myelinated axons (MA), yellow arrows, and synapses (blue circles) appear normal.
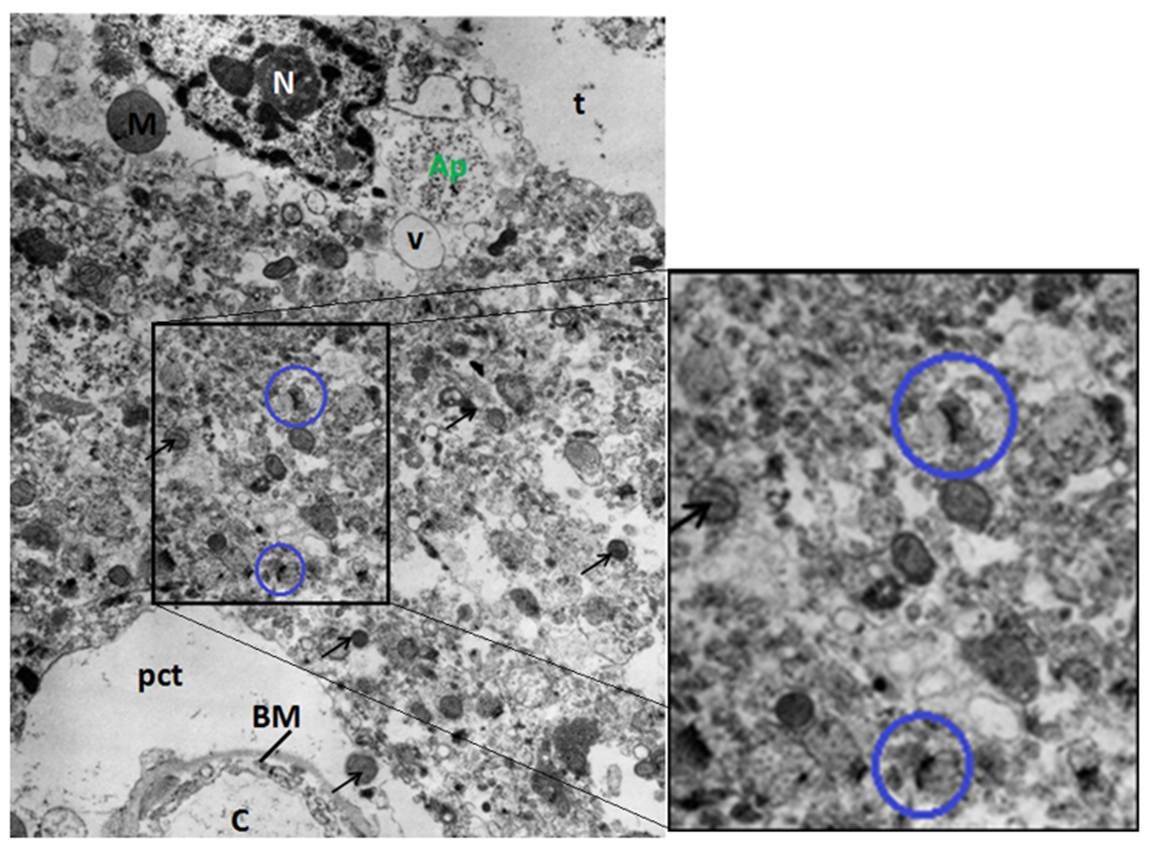
Figure 5: FIG-3, Cerebral Cortex, original magnification 15,000x. Typical disruption associated with freezing in the presence of ischemia and 3M glycerol is present; pericapillary (pct) and neuropil tears (t), as well as vacuolization (V) and debris. Despite this disruption, two synapses are visible (blue circles).
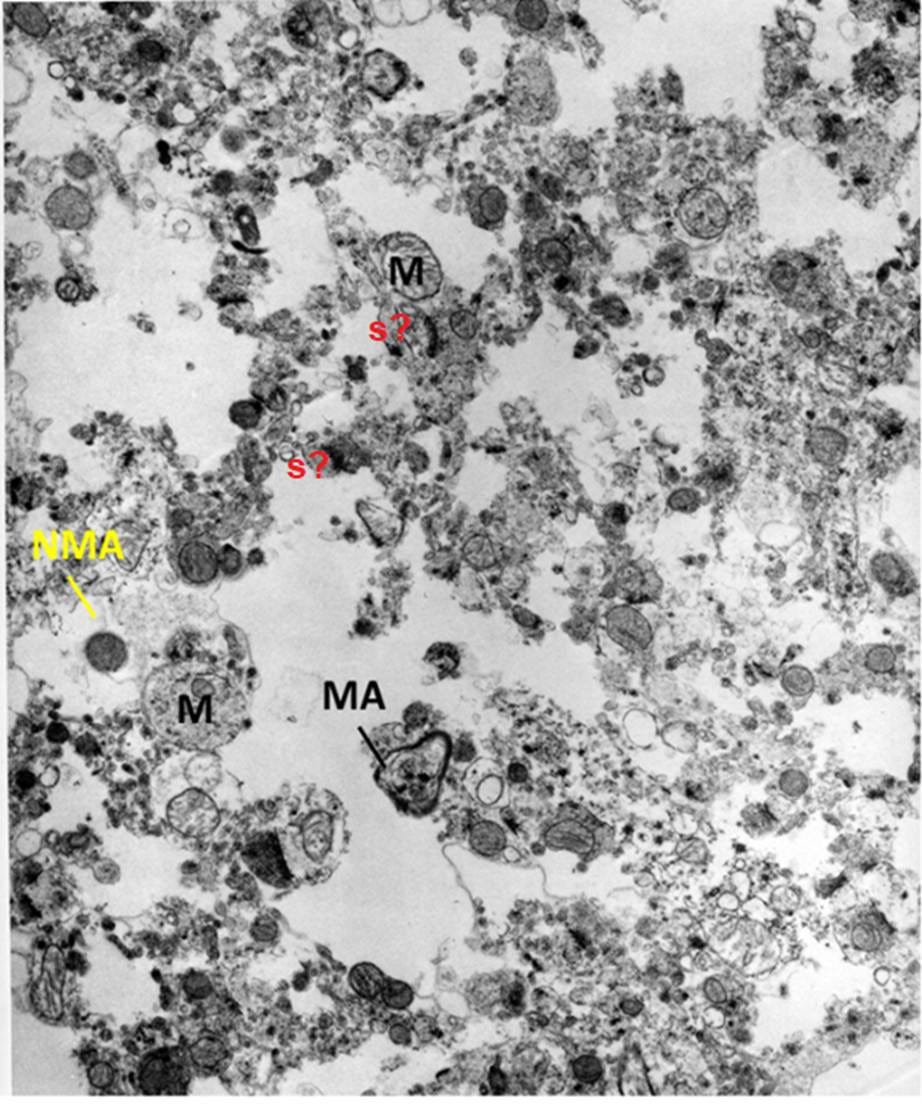
Figure 6: FIG-3, Cerebral Cortex, original magnification 15,000x. Nearly uniform maceration of the neuropil is reminiscent of a brain tissue homogenate. Some myelinated (MA) and non-myelinated axons (NMA) are visible, as is a very swollen mitochondrion (M), which retains what appears to be the debris of the cristae structure. There may be synapses (s?) present which appear as free-floating structures.
These two micrographs capture the spectrum of injury observed in the study and exemplify the nature and extent of the injury many cryonics patients likely experience today.
Another way to explain this is that the neuropil is likely subjected to multiple microscopic extrusion presses that transform some cells into amorphous debris. This is consistent with what we observe after thawing tissues that were frozen with grossly inadequate or absent cryoprotection. Because these forces and ice lattices are heterogeneous, this also explains why there are sometimes islands of comparatively recognizable tissue embedded in widespread, catastrophic destruction, which I’ve previously described as “debris” (Figure 6).
A closer analogy is to imagine information written on innumerable thin sheets of cold butter and then forcing those sheets through a coarse metal sieve under pressure. Fragments of the writing would survive, and some regions might remain surprisingly intact, but much of the original spatial configuration would be transformed into a heterogeneous mixture of compressed cell components and scattered debris.
The Nature of Cryoinjury
It is difficult to understand how such extensive tissue disruption can occur until the magnitude of the forces generated during ice growth is appreciated. In recent years, researchers in fields far removed from cryobiology—including geology, soil science, materials science, and concrete engineering—have begun reexamining the physical mechanisms responsible for frost heave, ice-induced fracture, and the destruction of porous materials during freezing (Baccile et al., 2020; Gerber et al., 2022; Webber & Worster, 2025). Using techniques such as synchrotron Small Angle X-ray Scattering (SAXS) and related methods, investigators have estimated the pressures generated and within the confined spaces between advancing ice fronts.
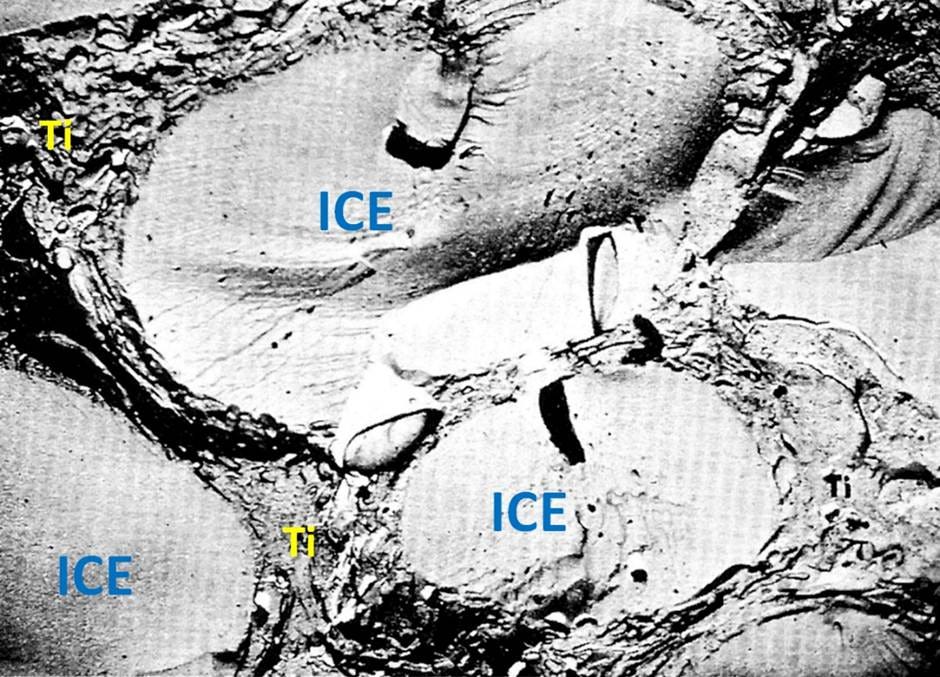
Figure 7. A freeze-fracture electron micrograph showing extracellular ice masses with the tissue architecture compacted between them. This intense compaction makes it impossible to image the tissue’s fine structure using conventional light microscopy or transmission electron microscopy.
The results are sobering. In one study, the pressure between growing ice crystals was estimated at approximately 1 kbar (14,500 psi; 987 atm) at −15 °C, increasing to 3.5 kbar (50,800 psi; 3,454 atm) at −60 °C (Baccile et al., 2020). Even allowing for methodological uncertainties, these values correspond to tens of thousands of pounds per square inch and thousands of atmospheres, illustrating the extreme mechanical forces exerted on material trapped between ice lattices.
It is important to emphasize that the model systems used by these investigators are not directly applicable to biological tissues. Their principal relevance to cryobiology lies in providing an order-of-magnitude estimate of the pressures that can be generated during ice growth and, to a lesser extent, in demonstrating that water within porous materials may be transported considerable distances through the matrix toward initial ice nucleation sites, where large, mechanically disruptive ice masses can develop. In biological tissues, however, the presence of cells embedded within a complex extracellular matrix imposes substantial structural and kinetic constraints on water transport over distances of micrometers, let alone centimeters.
Nevertheless, as Figure 8 illustrates, a substantial redistribution of water must occur over distances on the order of 5–10 µm. The resulting pattern indicates that ice formation in this brain is both anisotropic and a gross histological phenomenon, rather than a process confined to small crystals or ice masses immediately adjacent to the cells from which water was osmotically extracted. This morphology is consistent with the formation of large extracellular ice structures that reorganize tissue architecture across dimensions. It is also important to recognize that the size, morphology, and spatial distribution of ice within biological tissues are influenced by additional factors, including cooling rate and the extent of ice recrystallization during cooling to solidification.
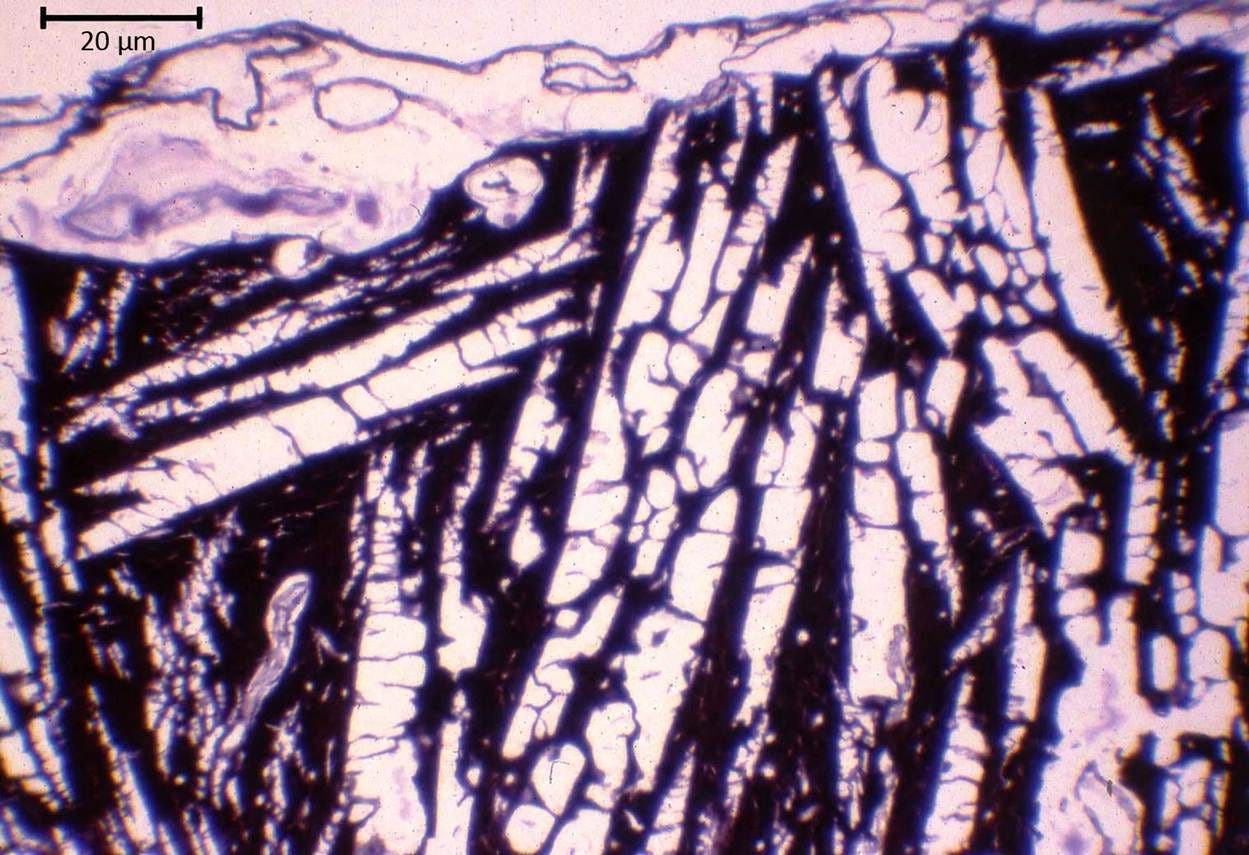
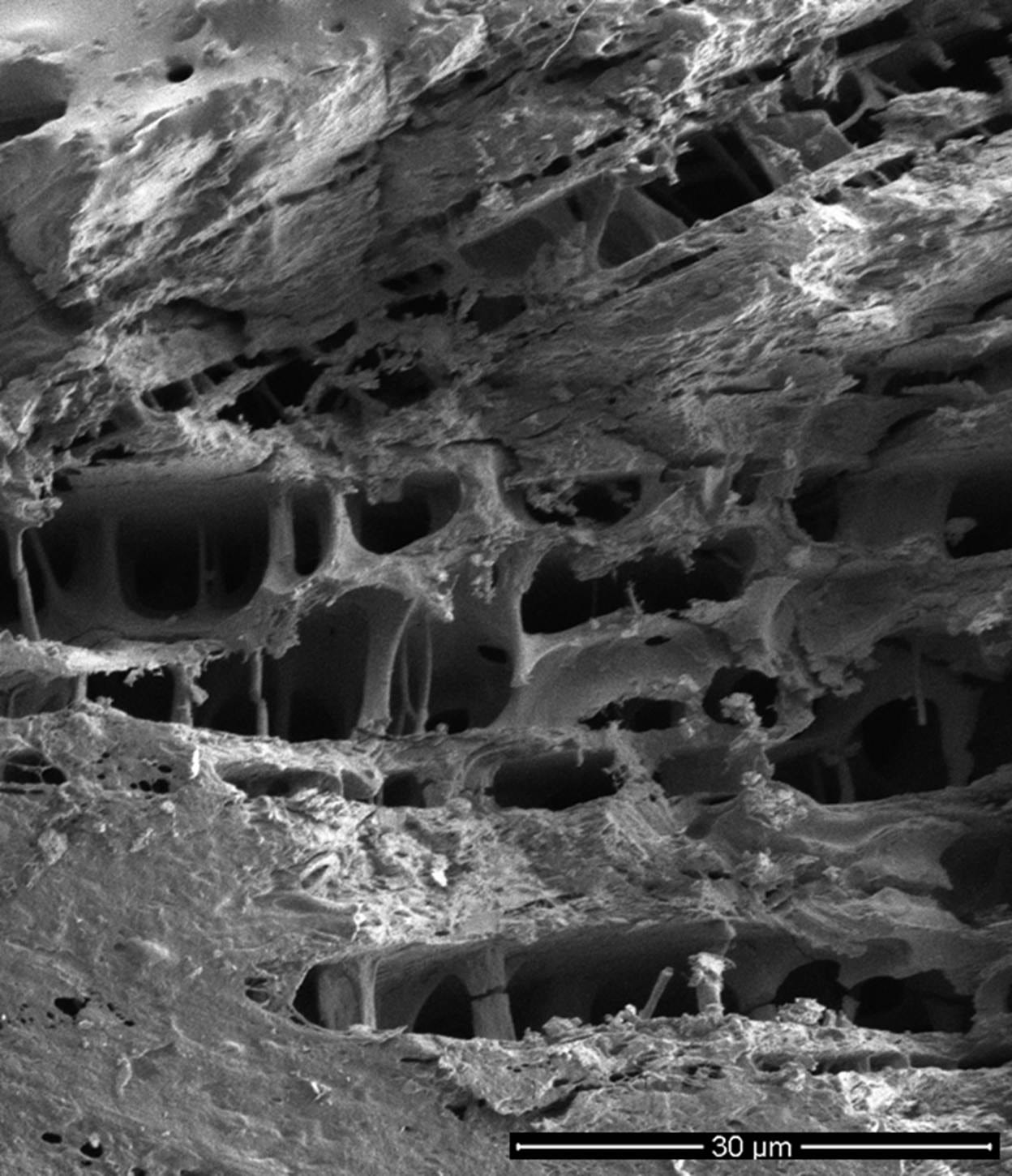
Figure 8. Imaging the location of ice in freeze-substituted rabbit cerebral cortex after equilibration with 3.72 and 3.0 M glycerol, and frozen to -80 ℃.
Top: Light micrograph (400×) of rabbit cerebral cortex after freezing and freeze-substitution following cryoprotection with 3.72 M glycerol. The large, pale spaces correspond to regions formerly occupied by extracellular ice, which was subsequently removed during freeze-substitution, while the darker, compressed bands contain the dehydrated, concentrated cellular elements of the neuropil. The structure atop the neuropil is the cerebral meninges. The image illustrates the extensive mechanical reorganization of brain tissue resulting from anisotropic growth of extracellular ice and suggests a complex three-dimensional architecture formed by multiple interacting ice fronts.
Bottom: Freeze-substituted and fractured rabbit cerebral cortex equilibrated with 3.0 M glycerol imaged using scanning electron microscopy (SEM) at 2000x magnification, revealing 3-dimensional detail not visible in the 2-dimensional tissue section in the top image. This image suggests, but cannot confirm, that the tissue’s membrane structure and connectivity are intact, albeit extensively distorted by ice. The rough and ragged surfaces are artifacts of fracturing the specimen from the original tissue block for imaging while it is dry and fragile. Similarly, the breaks in the interior columnar structures are likely due to specimen sectioning. What is striking about this image is that it appears to show the intersection of at least three discrete ice crystal planes.
See Appendix A for how the scale bar in the top image was approximated. These images are courtesy of Gregory M. Fahy, Ph.D., F.S.C., F.A.A.A., Executive Director and Chief Scientific Officer, 21st Century Medicine, Inc. Dr. Fahy conducted the research that produced this micrograph during the 1980s. The opinions and conclusions about this micrograph in this article are wholly those of the author and do not necessarily reflect those of Dr. Fahy.
Perspectives in cryonics are often shaped by engineering, computing, or everyday practical experiences, rather than formal training in biology. There is an implicit assumption that cells are miniature machines assembled from rigid parts. The problem is that cells, tissues, organs, and especially the brain, do not operate by those rules but instead obey the laws of soft-matter physics. A good antidote to this perspective is the excellent, highly accessible book The Machinery of Life (Goodsell, 2009), which provides detailed visual insight into what life is like at the nanoscale. The full text is available at no charge by clicking on the link embedded in the book’s title here.
Cells are not like the components of a computer, a cell phone, or an automobile. They are more akin to soap bubbles, with information encoded in their membranes and in a similarly plastic, reformable material inside, supported by a dynamic skeleton of proteins that undergo rapid, near-complete disassembly of their microtubule network into free tubulin subunits within minutes when cells are cooled to 0–5 °C (Brinkley & Cartwright, 1975; Hall, Evans & Hawortth, 1993; Desai & Mitichison, 1997). Indeed, I find it amazing that animals, including dogs, can completely recover from 4–5 hours of exposure to these temperatures. So, there is the danger, to use another analogy, that cryonicists are predicting the world’s behavior using Newtonian physics without considering relativity and quantum mechanics.
The root of the problem is that we do not know the condition of brain tissue in its frozen state before the forces of both freezing and thawing supervene. Thus, it is possible to argue that the catastrophic and likely information-destroying effects observed in thawed tissue are an artifact of thawing and do not reflect the tissue’s condition in the frozen state. Cryonics flourishes where data does not exist. Almost invariably, where there is ambiguity or uncertainty, optimism triumphs. And optimism is all too often the solvent of inquiry and validating experimentation.
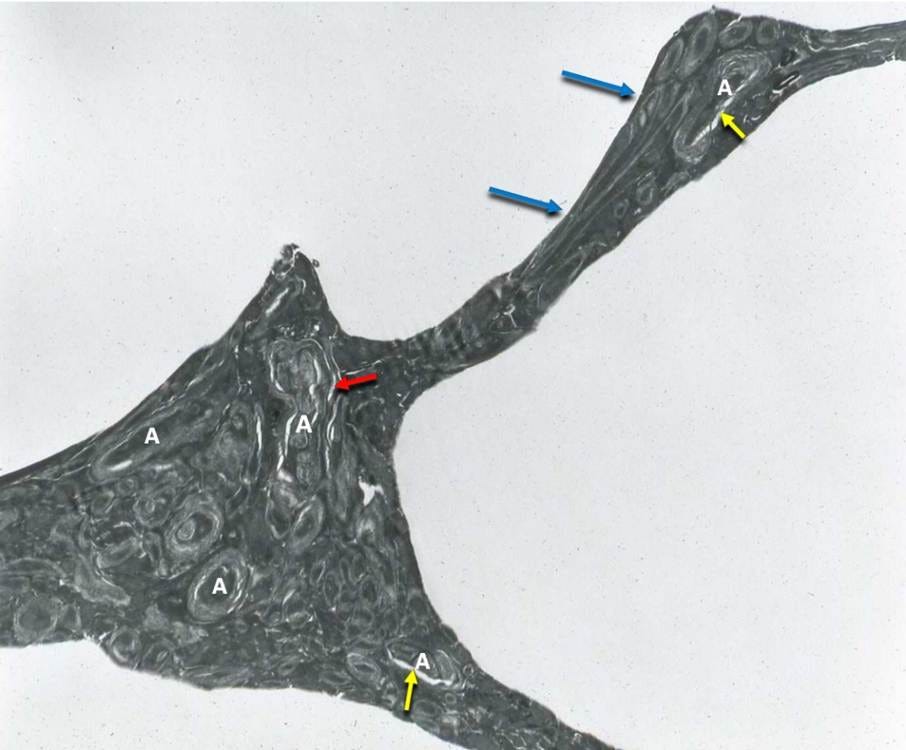
Figure 9. Freeze-substituted rabbit cerebral cortex perfused with 3.72 M Glycerol, original magnification 8,000×. The neuropil, consisting primarily of axons (A), is dehydrated and compressed between ice, with the tissue being stretched and deformed (blue arrows), indicating substantial elasticity during the interval when ice formed. The yellow arrows indicate dehydration of the axoplasm and its separation from the myelin, as is seen during glycerol perfusion without freezing. The red arrow indicates an area of myelin decompaction. This image is courtesy of Gregory M. Fahy, Ph.D., F.S.C., F.A.A.A., Executive Director and Chief Scientific Officer, 21st Century Medicine, Inc. Dr. Fahy conducted the research that produced this micrograph during the 1980s. The opinions and conclusions about this micrograph in this article are wholly those of the author and do not necessarily reflect those of Dr. Fahy.
What You Can’t See Can Hurt You
Of course, the problem is that we cannot directly image uncryoprotected frozen tissue using either light or electron microscopy because, as seen in Figure 7, it is dehydrated and compressed between extracellular ice masses. If a significant amount of cryoprotectant is present, the tissue can be examined by freeze-substitution, a process in which ice is dissolved and removed at ≈ −80 °C using an organic solvent containing fixatives and stains. When this is done with brains cryoprotected with 3 to 3.72 M glycerol, as shown in Figures 8 and 9, the dehydrated and compressed tissue appears to retain substantial structure, and the neuropil shows considerable elastic deformation. Remarkably, it is even possible to observe artifacts typically caused by glycerol perfusion itself, including dehydration and separation of axoplasm from myelin, as well as myelin decompaction associated with osmotic injury. Figure 8 (bottom) is particularly suggestive because it appears to show that ice may not sever intercellular connections, as seen in Figure 9, but may instead stretch and elastically deform the tissue without severing them. However, the resulting images do not provide sufficient resolution to determine whether cell membranes, synapses, or the connectome remain intact. In the absence of multimolar cryoprotection, even the limited structural detail visible in Figure 9 is not resolvable.
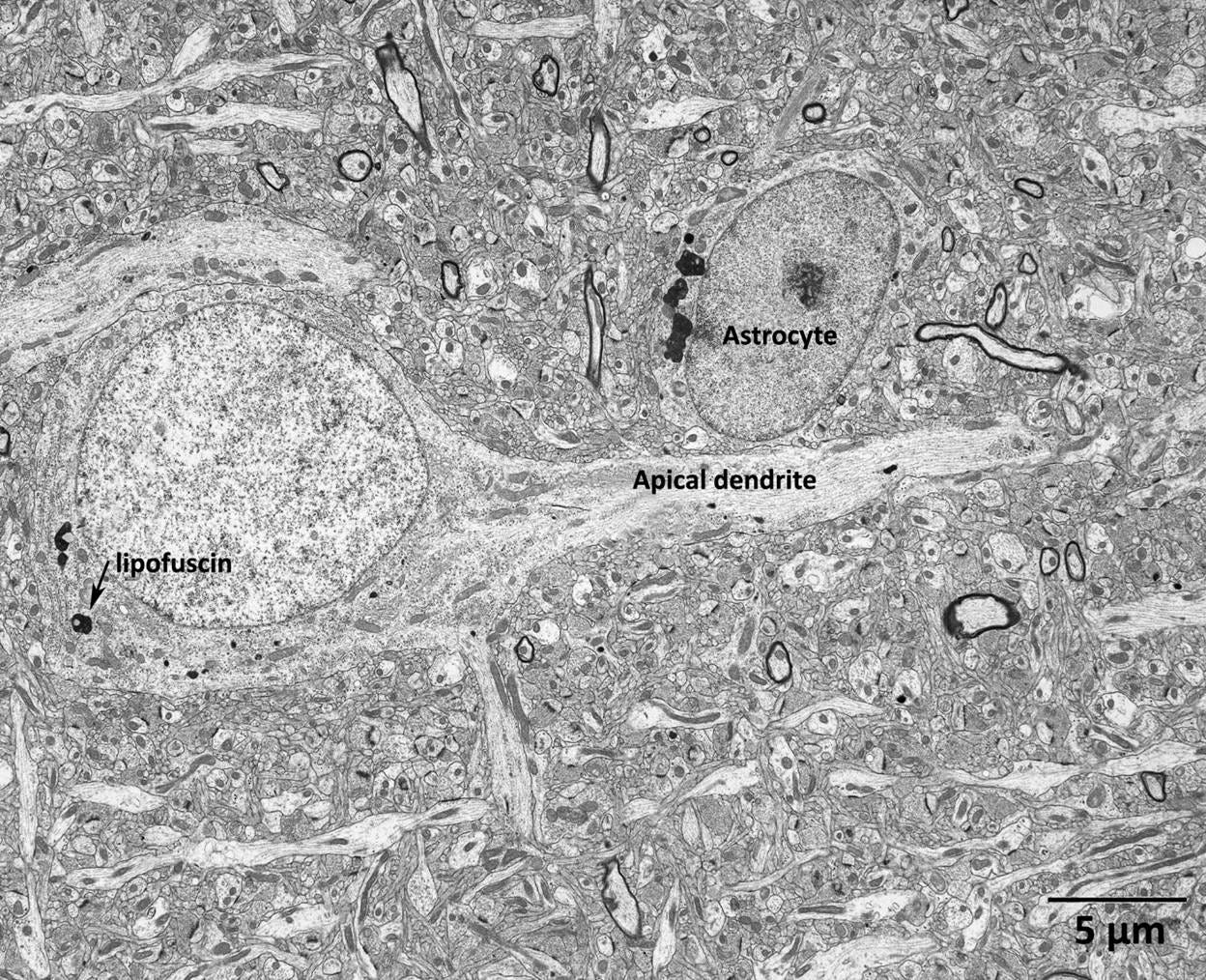
Figure 10. A pyramidal cell in layer 3 of the prefrontal cortex in a 5-year-old macaque monkey. This image is provided for visual comparison with the post-thaw micrograph of the rabbit cerebral cortex neuropil in Figure 9 below. The contrast between these images illustrates the profound structural disruption caused by cryoprotection, freezing, and thawing. The arrow indicates a lipofuscin granule. This annotation is an artifact of reusing the micrograph from a different context and is not significant for the present analysis. Image reproduced from (Peters, A., & Sethares, n.d.).
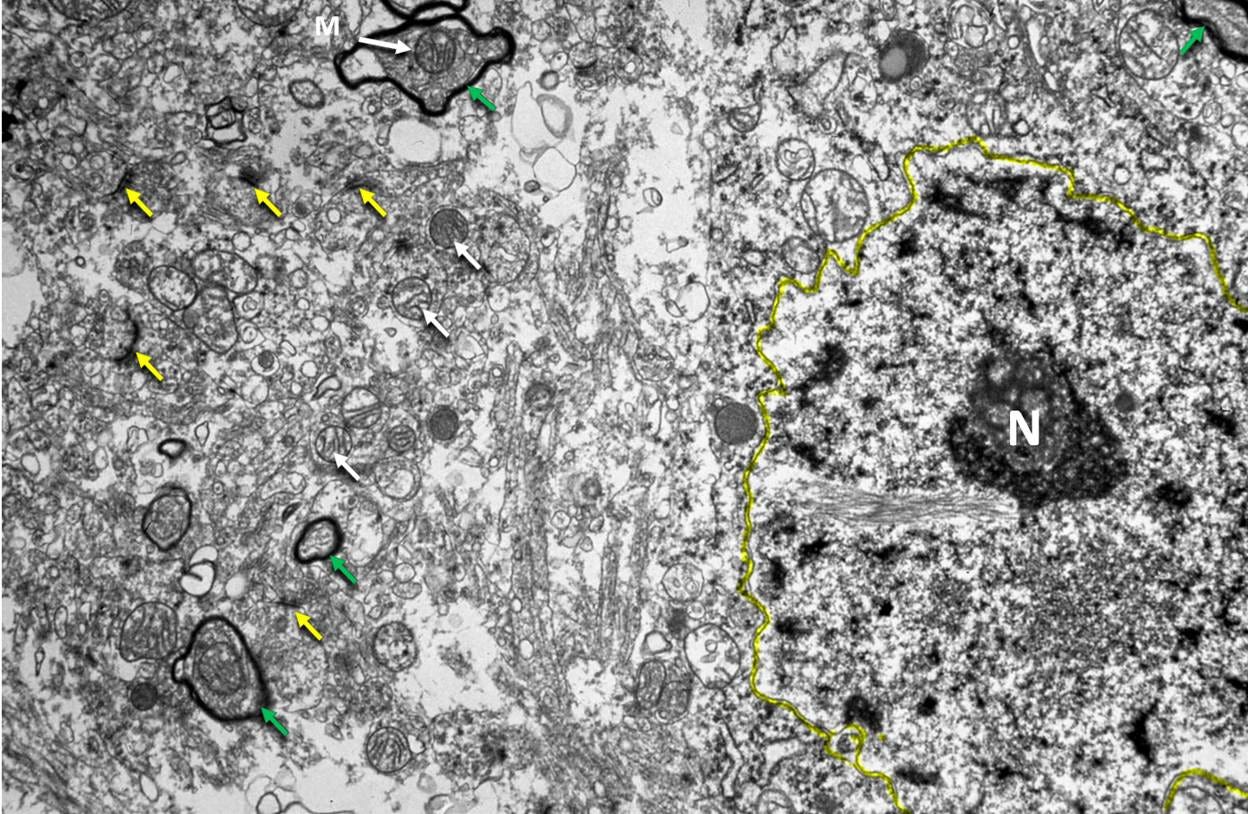
Figure 11. Rabbit cerebral cortex after perfusion with 3 M glycerol sat 20 ℃, freezing to -80 ℃, rewarming and reperfusion with aldehyde fixatives. Original magnification 7,000×. Synaptic clefts (yellow arrows), mitochondria (white arrows), myelinated axons (green arrows), a cell nucleus, and the plasma membrane of a cell (outlined in yellow) are visible, but have experienced significant degradation. Much of the left side of the image consists of open spaces and what appear to be macerated cellular debris. This image is courtesy of Gregory M. Fahy, Ph.D., F.S.C., F.A.A.A., Executive Director and Chief Scientific Officer, 21st Century Medicine, Inc. Dr. Fahy conducted the research that produced this micrograph during the 1980s. The opinions and conclusions about this micrograph in this article are wholly those of the author and do not necessarily reflect those of Dr. Fahy.
New Imaging Technologies Offer the Potential for New Insights
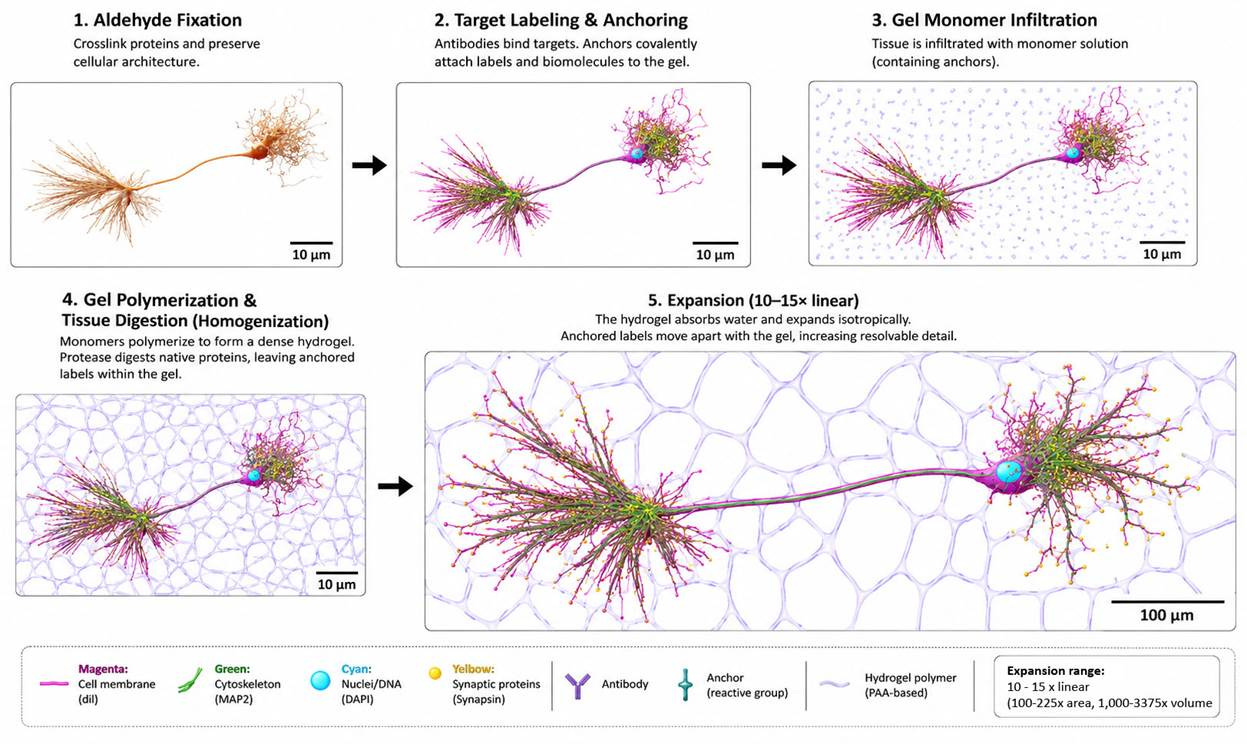
Figure 12. Procedure for expansion microscopy (ExM). Tissue is first fixed with aldehydes to preserve cellular architecture and then labeled with fluorescent probes, antibodies, lectins, or other molecular markers. Reactive anchoring molecules covalently link the labeled targets, or the biomolecules themselves, to a swellable hydrogel precursor solution that permeates the specimen. Following hydrogel polymerization, the tissue is mechanically homogenized, typically by proteolytic digestion or detergent-assisted denaturation, thereby allowing the polymer network to expand isotropically. Immersion in water causes the hydrogel to swell, increasing the linear dimensions of the specimen by approximately 10–15× (protocol dependent) while preserving the relative spatial relationships of the anchored targets. The resulting physical separation of cellular structures enables nanoscale imaging using conventional fluorescence microscopy. In principle, analogous approaches may be applicable to the study of dehydrated and mechanically compressed brain tissue, provided it is appropriately rehydrated and processed.
Expansion Microscopy and the Chrysalis Concept
Introduced in 2015, expansion microscopy (ExM) (Figure 12) bears a striking conceptual resemblance to Thomas Donaldson’s Chrysalis repair system (Donaldson, 1988). Donaldson argued that repair might be limited by the lack of space within the brain’s densely packed molecular structure. His solution was to expand the tissue isotropically, creating access for repair systems.
In ExM, the fixed tissue is first treated with bifunctional anchoring molecules that bind to target structures at one end and present a polymerizable “handle” at the other. The tissue is then infiltrated with a hydrogel monomer solution, which is polymerized into a dense, three-dimensionally cross-linked matrix that incorporates the anchored targets. Following polymerization, the specimen is treated with proteases that digest the cytoskeleton, extracellular matrix, and other proteins that hold cells and tissues together. This step removes the protein framework that would otherwise prevent expansion while leaving the anchored targets attached to the gel matrix. The hydrogel is then immersed in deionized water, causing it to expand uniformly in all three dimensions (Gallagher & Zhao, 2021). The desired amount of expansion separates the anchored targets, typically by approximately 4× to 15× linearly, allowing their architecture to be interrogated using conventional light microscopy (Gallagher & Zhao, 2021; Gambarotto et al., 2018).
“In such a situation, a chrysalis would first envelop the patient, then send tendrils of itself in between all of his cells. It disassembles the patient, surrounding each cell with its own repair machinery and vascular system. The disassembly process carefully preserves information about locations of the patient’s cells and their relationships to each other. If necessary, though, morphogen chemical gradients could also retain this information. A patient would swell up to ten times his original volume. After repair, the chrysalis slowly withdraws the same way it entered, re-establishing normal cell-to-cell relationships.”
—Thomas Donaldson, 24th Century Medicine
In the case of severely compacted tissue in the frozen state, the process would begin with freeze-substitution at ≈ -80 °C using an organic solvent such as acetone or methanol to dissolve the ice and remove residual tissue water. The solvent used during freeze-substitution would also contain aldehyde fixatives that cross-link the tissue in its frozen configuration. The tissue would then be transitioned into an aqueous environment, allowing the anchoring and monomer polymerization steps to proceed according to the standard protocol.
In conventional cryobiology and neuroscience, there has been no reason to date to apply such methods to frozen brain tissue. Only biostasis raises the question of whether severely dehydrated and compressed tissue retains the ultrastructural information that encodes memory and identity. Nothing in what I have written should imply that using ExM for this purpose will be straightforward. Nevertheless, the technique may enable determination of whether tissue compressed between ice masses retains cell membranes, intracellular architecture, and synaptic connectivity, or whether that information has been disrupted and dispersed to the point of irrecoverability.
It is here that we return to the problem of reading the Herculaneum scrolls, or, for that matter, similar problems encountered in deciphering some of the Dead Sea Scrolls (Anava et al., 2020). The challenge may ultimately be less one of imaging than of interpretation. The results of expansion microscopy will likely yield images that require sophisticated machine-learning methods for analysis. Here, the reader is referred to the excellent Vesuvius Challenge webpage, which details the immense multidisciplinary effort required to decipher the Herculaneum scrolls. Still, we will not know how opaque or problematic the interpretation of ExM data from frozen tissue will be until the work is begun.
One of the most attractive aspects of ExM is that it is comparatively easy to perform and does not require the costly infrastructure associated with electron microscopy. It is not an easy technique to master, but once mastered, it enables high-throughput analysis of both structure and biochemistry via a priori and post hoc labeling with sophisticated stains and markers.
Cryo-Electron Tomography
Although I have considered using expansion microscopy to study compressed frozen tissue ever since Aurelia Song first suggested the possibility during a walkabout in San Francisco more than five years ago, Zuber and Lučić’s 2022 review, Neurons as a Model System for Cryo-Electron Tomography (Cryo-ET), prompted me to think more broadly about the problem. In particular, it raised the question of whether cryogenic tomography—especially cryogenic synchrotron nanotomography or related X-ray imaging modalities—could be used to visualize brain tissue compressed to 20–30% of its original volume in the frozen state, without extensive processing, staining, or labeling (Figure 13).
Cryo-ET can directly visualize neurons and their internal architecture, including membranes, mitochondria, endoplasmic reticulum, cytoskeletal elements, synaptic vesicles, and other subcellular structures, without fixation, dehydration, staining, or embedding (Hylton & Swulius, 2021). Figure 13 shows a cryo-electron tomographic reconstruction of a presynaptic bouton, illustrating the level of ultrastructural detail that can be recovered from flash-vitrified neural tissue (Zuber & Lučić, 2022).
The principal limitation of Cryo-ET is that it can only be applied to extremely thin specimens (≈ 300–500 nm) (Baumeister, 2005; Oikonomou & Jensen, 2017; Hylton & Swulius, 2021). While this makes it a powerful tool for studying cellular ultrastructure, it cannot presently be used to image the volumes of tissue required to determine the topology of cell membranes compressed within frozen tissue, much less large-scale neuronal connectivity.
Synchrotron Nanotomography as a Path Forward
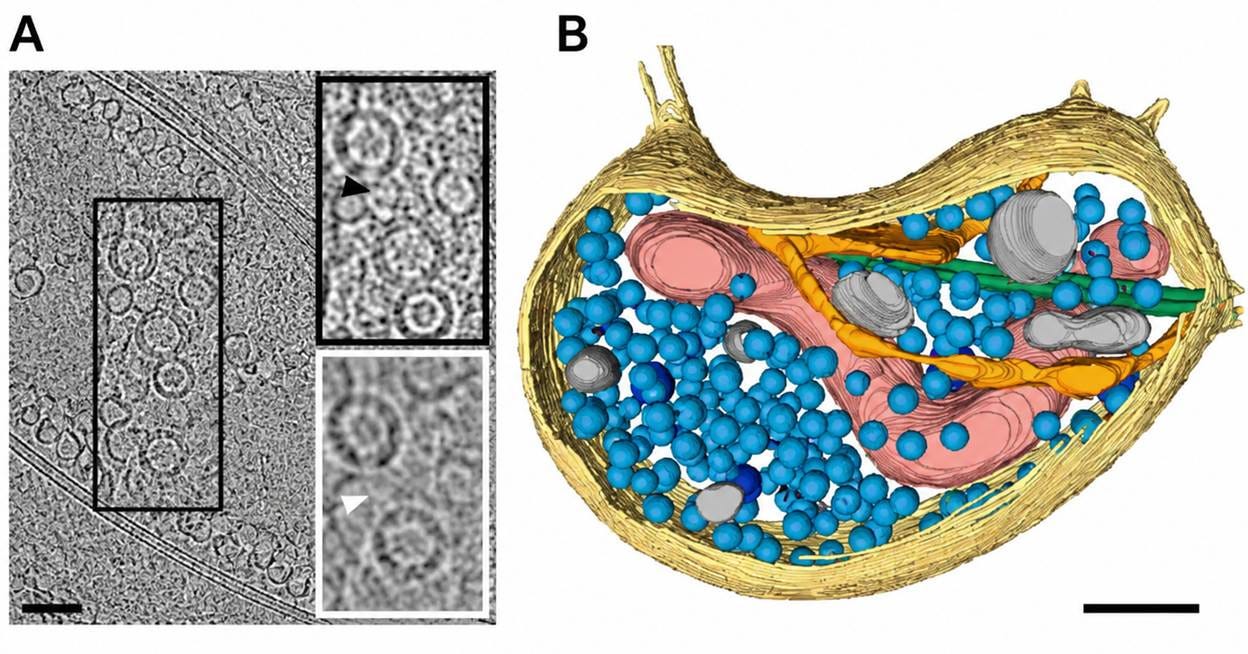
Figure 13. Neuron cell-ubiquitous structures observed by Cryo-ET. (A) Low-magnification cryo-ET slice of a synaptic bouton from a dissociated neuronal culture. The boxed region is magnified to the right: a synaptic vesicle (black arrowhead) and a dense core vesicle (white arrowhead) are indicated. (B) Three-dimensional segmentation of the bouton in A, showing plasma membrane (yellow), mitochondria (red), microtubules (green), smooth ER (orange), synaptic vesicles (light blue), dense core vesicles (dark blue), and other membranous compartments (grey). Scale bars: (A) 100 nm; insets 50 nm; (B) 100 nm. Reproduced from Zuber and Lučić (2022).
The work of Kuan et al. (2020) and Bosch et al. (2025) has made this possibility more compelling. Their studies demonstrate that X-ray nanotomography and related approaches, including ptychographic X-ray tomography (PXCT), a lensless coherent imaging method capable of nanoscale three-dimensional reconstruction, can non-destructively resolve neuronal and subcellular architecture within tissue volumes far larger than those accessible to Cryo-ET.
Thus, it is conceivable that cryogenic synchrotron nanotomography, ptychographic X-ray tomography (PXCT), or a successor technology in that family could eventually permit direct imaging of dehydrated, compacted brain tissue while it remains frozen, eliminating the need for thawing, freeze-substitution, or other processing steps that introduce artifacts. Existing synchrotron-based imaging technologies have already demonstrated three-dimensional imaging of frozen-hydrated biological specimens (Diaz, 2015). The crucial unresolved question is whether sufficient resolution and contrast can be achieved to resolve membrane topology and synaptic architecture from tissue compressed between extracellular ice masses.
Conclusion: Knowing as a Priority
The likely state of patients subjected to extensive freezing is experimentally answerable. Doing so will require substantial resources, advances in imaging and computational reconstruction, and, most importantly, investigators willing to pursue a problem that conventional cryobiology has had no reason to address. Undertaking this work requires the same elements as the Vesuvius Challenge: funding, passion among multidisciplinary investigators, and a profound commitment to evaluating cryopreservation under adverse conditions. This final requirement is the hardest to meet. Although the history of cryonics has many profoundly adverse cryopreservations, no one—from management to membership—wants to admit that their own case might become one of them. The ideal is ever the enemy of the real.
Alcor’s review of cases from 2000–2020 found that ≈ 23% of patients received no cryoprotective perfusion and were frozen without cryoprotection. Only ≈ 43% of patients achieved the terminal cryoprotectant concentration considered necessary for vitrification, while 31% underwent cryoprotective perfusion but failed to reach that target (Benjamin & de Wolf, 2021a, 2021b). These cases frequently occur due to logistical constraints and the reality that death is not infrequently sudden and unexpected. As a consequence, there is little reason to expect that severely compromised cryopreservations will become rare in the foreseeable future.
I believe that conducting this research should be one of the highest priorities of biostasis organizations (i.e., cryonics organizations) that accept “all comers” –patients who present with a wide range of conditions that result in a similarly broad range of injuries. For this reason, I have repeatedly argued that biostasis should be restructured as a legitimate clinical trial, with clearly defined inclusion and exclusion criteria, in which only patients who meet predetermined clinical standards are accepted for care. Such eligibility criteria are a foundational feature of modern clinical research because they permit meaningful evaluation of outcomes, facilitate comparison among patients, and support the systematic refinement of treatment protocols over time (Pocock, 1983; Friedman et al., 2015). According to the Shandong Yinfeng Life Science Research Institute (n.d.), this is the paradigm they employ in operating its human cryopreservation program in China (Shandong Yinfeng Life Science Research Institute, n.d.). Similarly, based on multiple discussions with Borys Wróbel, Chief Scientist at Nectome, Inc., over the past six months (personal communications, 2025–2026), I understand that Nectome intends to employ a similar paradigm.
This would not preclude the continued practice of cryonics. People should be free to practice their religious and philosophical beliefs so long as they do not harm others and are adequately informed about the nature of the procedures used on them. And indeed, were it possible to demonstrate that brain tissue in suboptimally treated patients still contained sufficient structural information to reasonably support a case for recovery, it would go a long way toward moving the practice from ritualistic optimism to fact-based science.
References
Anava, S., Neuhof, M., Gingold, H., Sagy, O., Munters, A., Svensson, E. M., Afshinnekoo, E., Danko, D., Foox, J., Shor, P., Riestra, B., Huchon, D., Mason, C. E., Mizrahi, N., Jakobsson, M., & Rechavi, O. (2020). Illuminating Genetic Mysteries of the Dead Sea Scrolls. Cell, 181(6), 1218-1231.e27. https://doi.org/10.1016/j.cell.2020.04.046
Baccile, N., Zinn, T., Laurent, G., Messaoud, G., Cristiglio, V., & Fernandes, F. (2020, February 26). Unveiling the Interstitial Pressure Between Growing Ice Crystals During Ice-Templating Using a Lipid Lamellar Probe. arXiv.Org. https://doi.org/10.1021/acs.jpclett.9b03347
Baldwin, C., Sullivan, M., & Drake, A. (2013, August). Alcor A-1088 case report. Alcor Life Extension Foundation. Archived by the Internet Archive Wayback Machine. https://web.archive.org/web/20131115000000/http://alcor.org/Library/pdfs/casereportA1088DennisRoss.pdf
Benjamin, M., & de Wolf, A. (2021a). Alcor case metrics 1967–1999. Cryonics, 42(3), 12–21. Retrieved from https://www.cryonicsarchive.org/docs/cryonics-magazine-2021-03.pdf
Benjamin, M., & de Wolf, A. (2021b). Alcor case metrics 1967–2020. Cryonics, 43(4), 28–31. Retrieved from https://www.cryonicsarchive.org/docs/cryonics-magazine-2021-04.pdf
Bosch, C., Aidukas, T., Holler, M., Pacureanu, A., Müller, E., Peddie, C. J., Zhang, Y., Cook, P., Collinson, L., Bunk, O., Menzel, A., Guizar-Sicairos, M., Aeppli, G., Diaz, A., Wanner, A. A., & Schaefer, A. T. (2025). Nondestructive X-ray tomography of brain tissue ultrastructure. Nature Methods, 22(12), 2631–2638. https://doi.org/10.1038/s41592-025-02891-0
Baumeister, Wolfgang (2005). Electron tomography: Towards visualizing the molecular organization of the cytoplasm. Current Opinion in Structural Biology, 15(2), 225–232.
Brinkley, B. R., & Cartwright, J. (1975). Cold-labile and cold-stable microtubules in the mitotic spindle of mammalian cells. Annals of the New York Academy of Sciences, 253, 428–439. https://doi.org/10.1111/j.1749-6632.1975.tb19218.x
Ceccarelli, S., Rippa, M., Caruso, G., Luvidi, L., Boccuti, S., Paturzo, M., Pagliarulo, V., Fleischer, K., Miliani, C., & Ranocchia, G. (2025). Pulsed thermographic analysis of Herculaneum papyri. Scientific Reports, 15(1), 34466. https://doi.org/10.1038/s41598-025-19911-w
Desai, A., & Mitchison, T. J. (1997). MICROTUBULE POLYMERIZATION DYNAMICS. Annual Review of Cell and Developmental Biology, 13(Volume 13, 1997), 83–117. https://doi.org/10.1146/annurev.cellbio.13.1.83
Diaz, A., Malkova, B., Holler, M., Guizar-Sicairos, M., Lima, E., Panneels, V., Pigino, G., Bittermann, A. G., Wettstein, L., Tomizaki, T., Bunk, O., Schertler, G., Ishikawa, T., Wepf, R., & Menzel, A. (2015). Three-dimensional mass density mapping of cellular ultrastructure by ptychographic X-ray nanotomography. Journal of Structural Biology, 192(3), 461–469. https://doi.org/10.1016/j.jsb.2015.10.008
Donaldson, T. K. (1988, December). 24th century medicine. Cryonics, 9(12), 16–34. Retrieved November 22, 2025, from Cryonics Archive PDF
Friedman, L. M., Furberg, C. D., DeMets, D. L., Reboussin, D. M., & Granger, C. B. (2015). Fundamentals of Clinical Trials. Springer.
Gallagher, B. R., & Zhao, Y. (2021). Expansion microscopy: A powerful nanoscale imaging tool for neuroscientists. Neurobiology of Disease. https://doi.org/10.1016/j.nbd.2021.105362
Gambarotto, D., Zwettler, F. U., Le Guennec, M., Schmidt-Cernohorska, M., & Et., A. (2018). Imaging cellular ultrastructures using expansion microscopy (U-ExM). Nature Methods. https://doi.org/10.1038/s41592-018-0238-1
Gerber, D., Wilen, L. A., Poydenot, F., Dufresne, E. R., & Style, R. W. (2022). Stress accumulation by confined ice in a temperature gradient. Proceedings of the National Academy of Sciences, 119(31), e2200748119. https://doi.org/10.1073/pnas.2200748119
Goodsell, D. S. (2009). The machinery of life (2nd ed., corrected). Copernicus Books.
Hall, S. M., Evans, J., & Haworth, S. G. (1993). Influence of cold preservation on the cytoskeleton of cultured pulmonary arterial endothelial cells. American Journal of Respiratory Cell and Molecular Biology, 9(1), 106–114. https://doi.org/10.1165/ajrcmb/9.1.106
Hylton, R. K., & Swulius, M. T. (2021). Challenges and triumphs in cryo-electron tomography. iScience, 24(9), 102959. https://doi.org/10.1016/j.isci.2021.102959
Kuan, A. T., Phelps, J. S., Thomas, L. A., Nguyen, T. M., Han, J., Chen, C.-L., Azevedo, A. W., Tuthill, J. C., Funke, J., Cloetens, P., Pacureanu, A., & Lee, W.-C. A. (2020). Dense neuronal reconstruction through X-ray holographic nanotomography. Nature Neuroscience, 23(12), 1637–1643. https://doi.org/10.1038/s41593-020-0704-9
Parsons, S., Parker, C. S., Chapman, C., Hayashida, M., & Seales, W. B. (2023). EduceLab-Scrolls: Verifiable Recovery of Text from Herculaneum Papyri using X-ray CT.
Peters, A., & Sethares, C. F. (n.d.). Chapter 1: Neuronal cell bodies. The Fine Structure of the Aging Brain. Boston University School of Medicine. Retrieved June 8, 2026, from The Fine Structure of the Aging Brain – Chapter 1: Neuronal Cell Bodies
Pichugin, Y., Fahy, G. M., & Morin, R. (2006). Cryopreservation of rat hippocampal slices by vitrification. Cryobiology, 52(2), 228–240. https://doi.org/10.1016/j.cryobiol.2005.11.006
Pocock, S. J. (1983). Clinical trials: A practical approach. John Wiley & Sons.
Radomski, C. (2026, May 28). Reading the unreadable: From the Herculaneum scrolls to the cryopreserved brain. The Biostasis Standard.

Shandong Yinfeng Life Science Research Institute. (n.d.). Yinfeng Life Extension Program. Retrieved June 4, 2026, from https://en.yinfenglife.org.cn/jihuagaishu.html
Tillberg, P. W., & Chen, F. (2019). Expansion Microscopy: Scalable and Convenient Super-Resolution Microscopy. Annual Review of Cell and Developmental Biology. https://doi.org/10.1146/annurev-cellbio-100818-125320
Webber, J. J., & Grae Worster, M. (2025). Cryosuction and freezing hydrogels. Proceedings of the Royal Society A: Mathematical, Physical and Engineering Sciences, 481(2310), 20240721. https://doi.org/10.1098/rspa.2024.0721
Zuber, B., & Lučić, V. (2022). Neurons as a model system for cryo-electron tomography. Journal of Structural Biology: X, 6, 100067. https://doi.org/10.1016/j.yjsbx.2022.100067
Appendix A
The light micrograph shown in Figure 8 is an accurate digital reproduction of an original 35 mm color transparency and was reported to have been acquired at a nominal magnification of 400×. Because no scale bar was present on the original image, a retrospective estimate was derived from the reported magnification.
This estimate assumes that the image occupied the full width of the original 35 mm slide frame and that the reported magnification was accurate. Because the original calibration data are unavailable, the resulting scale bar should be regarded as approximate, with an estimated uncertainty of ±10–20%.
A standard 35 mm slide contains an image area approximately 36 mm wide by 24 mm high. At a nominal magnification of 400×, the specimen field is approximately 90 µm wide by 60 µm high. The scanned image used in this study has a width of 1536 pixels, yielding an estimated scale of 0.0586 µm per pixel (90 µm ÷ 1536 pixels).
Based on this calibration, the 2 µm scale bar shown in Figure 8 corresponds to approximately 34 pixels. While this estimate cannot substitute for direct calibration of the original microscope system, it provides a reasonable approximation of the dimensions of the structures visible in the image and allows comparison with other micrographs presented in this article.
Hi Mike, Carrie's last name is "Radomski", please correct!